TJP1 Antikörper
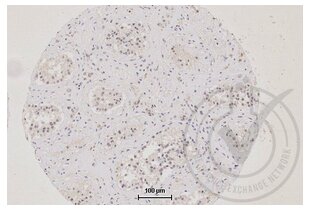
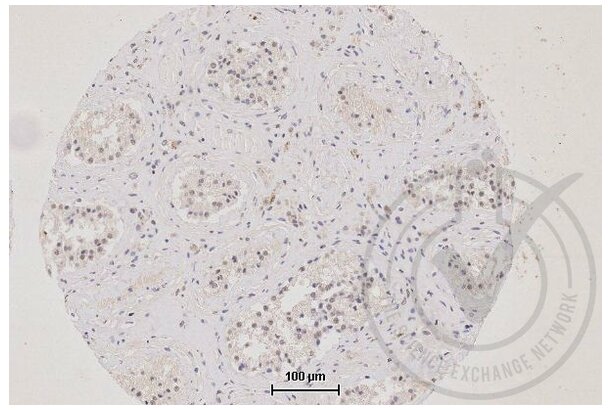 TJP1 Antikörper (AA 1551-1702) (ABIN675024)
TJP1 Antikörper (AA 1551-1702) (ABIN675024)
TJP1 Reaktivität: Human, Maus, Ratte, Schwein, Rind (Kuh) WB, ELISA, FACS, IHC (p), IF (p), IF (cc), IHC (fro) Wirt: Kaninchen Polyclonal unconjugated
TJP1 Reaktivität: Human, Maus, Ratte, Schwein, Rind (Kuh), Hund, Meerschweinchen, Kaninchen, Pferd, Affe, Huhn, Avian, Cat, Esel, Ziege, Hamster, Schaf WB, IF, IHC (p), IHC (fro) Wirt: Ziege Polyclonal unconjugated
KO Validated TJP1 Reaktivität: Human WB, IP, IF, ICC Wirt: Kaninchen Polyclonal unconjugated
TJP1 Antikörper nach Grade
Hier sind TJP1 Antikörper mit einem spezifischen Grade zu finden. Die hier aufgeführten Grade sind einige der verfügbaren. Ein Klick auf den entsprechenden Link führt zu den Produkten.
TJP1 Antikörper nach Wirt
Hier sind TJP1 Antikörper mit einem spezifischen Wirt zu finden. Die hier aufgeführten Wirt sind einige der verfügbaren. Ein Klick auf den entsprechenden Link führt zu den Produkten.
TJP1 Antikörper nach Klonalität
Finden Sie verfügbare monoklonale oder polyklonale TJP1 Antikörper. Klicken Sie auf einen Link, um zu den entsprechenden Produkten zu gelangen.
TJP1 Antikörper nach Klon
Hier sind TJP1 Antikörper mit einem spezifischen Klon zu finden. Die hier aufgeführten Klon sind einige der verfügbaren. Ein Klick auf den entsprechenden Link führt zu den Produkten.
Häufig verwendete TJP1 Antikörper
- (7)
- (7)
- (1)
- (2)
- (11)
- (1)
- (8)
- (7)
- (6)
- (1)
- (6)
- (1)
- (6)
- (2)
- (3)
- (4)
- (4)
- (4)
- (4)
- (3)
- (1)
- (2)
- (2)
- (2)
- (2)
Aktuelle Publikationen für unsere TJP1 Antikörper
: "Epithelial Heat Shock Proteins Mediate the Protective Effects of Limosilactobacillus reuteri in Dextran Sulfate Sodium-Induced Colitis." in: Frontiers in immunology, Vol. 13, pp. 865982, (2022) (PubMed).: "Pinocembrin alleviates ulcerative colitis in mice via regulating gut microbiota, suppressing TLR4/MD2/NF-κB pathway and promoting intestinal barrier." in: Bioscience reports, Vol. 40, Issue 7, (2021) (PubMed).
: "3D Organoid Culture From Adult Salivary Gland Tissues as an ex vivo Modeling of Salivary Gland Morphogenesis." in: Frontiers in cell and developmental biology, Vol. 9, pp. 698292, (2021) (PubMed).
: "Parasite-Derived Excretory-Secretory Products Alleviate Gut Microbiota Dysbiosis and Improve Cognitive Impairment Induced by a High-Fat Diet." in: Frontiers in immunology, Vol. 12, pp. 710513, (2021) (PubMed).
: "Psychoactive Effects of Lactobacillus johnsonii Against Restraint Stress-Induced Memory Dysfunction in Mice Through Modulating Intestinal Inflammation and permeability-a Study Based on the Gut-Brain ..." in: Frontiers in pharmacology, Vol. 12, pp. 662148, (2021) (PubMed).
: "Early-Life Intervention Using Exogenous Fecal Microbiota Alleviates Gut Injury and Reduce Inflammation Caused by Weaning Stress in Piglets." in: Frontiers in microbiology, Vol. 12, pp. 671683, (2021) (PubMed).
: "The Antimicrobial Peptide Mastoparan X Protects Against Enterohemorrhagic Escherichia coli O157:H7 Infection, Inhibits Inflammation, and Enhances the Intestinal Epithelial Barrier." in: Frontiers in microbiology, Vol. 12, pp. 644887, (2021) (PubMed).
: "Astragalus membranaceus and Salvia miltiorrhiza ameliorates cyclosporin A-induced chronic nephrotoxicity through the "gut-kidney axis"." in: Journal of ethnopharmacology, Vol. 269, pp. 113768, (2021) (PubMed).
: "Supplemental Bacillus subtilis DSM 29784 and enzymes, alone or in combination, as alternatives for antibiotics to improve growth performance, digestive enzyme activity, anti-oxidative status, immune ..." in: The British journal of nutrition, Vol. 125, Issue 5, pp. 494-507, (2021) (PubMed).
: "Zizyphus jujuba cv. Muzao polysaccharides enhance intestinal barrier function and improve the survival of septic mice." in: Journal of food biochemistry, Vol. 45, Issue 5, pp. e13722, (2021) (PubMed).
Aliase für TJP1 Antikörper
tight junction protein 1 (TJP1) Antikörpertight junction protein 1 (Tjp1) Antikörper
tight junction protein ZO-1 (LOC396567) Antikörper
ZO-1 Antikörper
ZO1 Antikörper
zo1 Antikörper
Haben Sie etwas anderes gesucht?
- TJAP1 Antikörper
- Titin Antikörper
- Tissue factor Antikörper
- TIRAP Antikörper
- TIPRL Antikörper
- TIPIN Antikörper
- Tiparp Antikörper
- TINF2 Antikörper
- TINAGL1 Antikörper
- TINAG Antikörper
- TIMP4 Antikörper
- TIMP3 Antikörper
- TIMP1 Antikörper
- TIMM9 Antikörper
- TIMM8B Antikörper
- TIMM8A/DDP Antikörper
- TIMM50 Antikörper
- TIMM44 Antikörper
- TIMM29 Antikörper
- TIMM23 Antikörper
- TJP2 Antikörper
- TJP3 Antikörper
- TK1 Antikörper
- TK2 Antikörper
- TKT Antikörper
- TKTL2 Antikörper
- TLE1 Antikörper
- TLE2 Antikörper
- TLE3 Antikörper
- TLE4 Antikörper
- TLE6 Antikörper
- TLK1 Antikörper
- TLK2 Antikörper
- TLL1 Antikörper
- TLN1 Antikörper
- TLN2 Antikörper
- TLR1 Antikörper
- TLR10 Antikörper
- TLR11 Antikörper
- TLR12 Antikörper



