GDNF Antikörper
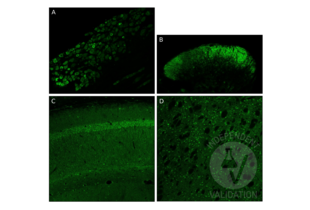
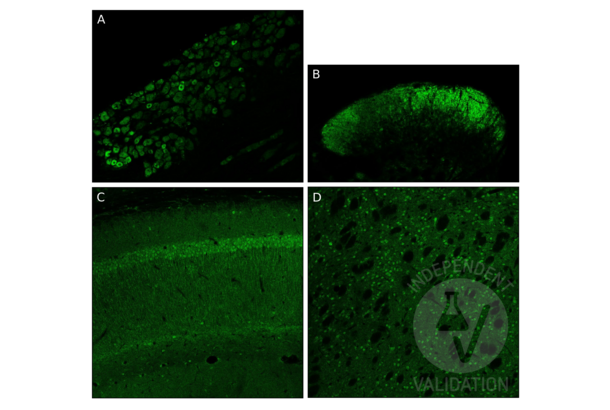 GDNF Antikörper (AA 121-211) (ABIN736536)
GDNF Antikörper (AA 121-211) (ABIN736536)
GDNF Reaktivität: Human, Maus, Ratte, Rind (Kuh), Kaninchen WB, ELISA, FACS, IF (cc), IF (p), IHC (p), IHC (fro) Wirt: Kaninchen Polyclonal unconjugated
GDNF Reaktivität: Human WB, IHC, ELISA, IP, FACS Wirt: Kaninchen Monoclonal 11D2 unconjugated Recombinant Antibody
GDNF Reaktivität: Human, Ratte WB, ICC, IHC (p) Wirt: Kaninchen Polyclonal unconjugated
GDNF Antikörper nach Reaktivität
Hier sind GDNF Antikörper für eine Vielzahl von Species wie anti-Human GDNF, anti-Mouse GDNF, anti-Rat GDNF zu finden. Die unten aufgeführten Species gehören zu den verfügbaren Arten. Klicken Sie auf einen Link, um zu den entsprechenden Produkten zu gelangen.
GDNF Antikörper nach Wirt
Hier sind GDNF Antikörper mit einem spezifischen Wirt zu finden. Die hier aufgeführten Wirt sind einige der verfügbaren. Ein Klick auf den entsprechenden Link führt zu den Produkten.
GDNF Antikörper nach Klonalität
Finden Sie verfügbare monoklonale oder polyklonale GDNF Antikörper. Klicken Sie auf einen Link, um zu den entsprechenden Produkten zu gelangen.
GDNF Antikörper nach Klon
Hier sind GDNF Antikörper mit einem spezifischen Klon zu finden. Die hier aufgeführten Klon sind einige der verfügbaren. Ein Klick auf den entsprechenden Link führt zu den Produkten.
GDNF Antikörper nach Fragment
Hier sind GDNF Antikörper mit einem spezifischen Fragment zu finden. Die hier aufgeführten Fragment sind einige der verfügbaren. Ein Klick auf den entsprechenden Link führt zu den Produkten.
Häufig verwendete GDNF Antikörper
- (1)
- (5)
- (1)
- (5)
- (4)
- (4)
- (5)
- (3)
- (3)
- (3)
- (3)
- (3)
- (3)
- (3)
- (3)
- (3)
- (3)
- (3)
Aktuelle Publikationen für unsere GDNF Antikörper
: "Co-overexpression of VEGF and GDNF in adipose-derived stem cells optimizes therapeutic effect in neurogenic erectile dysfunction model." in: Cell proliferation, Vol. 53, Issue 2, pp. e12756, (2020) (PubMed).: "Surface functionalization of TiO2 nanotubes with minocycline and its in vitro biological effects on Schwann cells." in: Biomedical engineering online, Vol. 17, Issue 1, pp. 88, (2018) (PubMed).
: "Non-invasive, neuron-specific gene therapy by focused ultrasound-induced blood-brain barrier opening in Parkinson's disease mouse model." in: Journal of controlled release : official journal of the Controlled Release Society, Vol. 235, pp. 72-81, (2016) (PubMed).
: "Normalization of ventral tegmental area structure following acupuncture in a rat model of heroin relapse." in: Neural regeneration research, Vol. 9, Issue 3, pp. 301-7, (2014) (PubMed).
: "Lipopolysaccharide inhibits the self-renewal of spermatogonial stem cells in vitro via downregulation of GDNF expression in Sertoli cells." in: Reproductive toxicology (Elmsford, N.Y.), Vol. 45, pp. 87-93, (2014) (PubMed).
: "Antenatal taurine reduces cerebral cell apoptosis in fetal rats with intrauterine growth restriction." in: Neural regeneration research, Vol. 8, Issue 23, pp. 2190-7, (2014) (PubMed).
: "S100B protein, brain-derived neurotrophic factor, and glial cell line-derived neurotrophic factor in human milk." in: PLoS ONE, Vol. 6, Issue 6, pp. e21663, (2011) (PubMed).
: "Glial cell line-derived neurotrophic factor is essential for postnatal survival of midbrain dopamine neurons." in: The Journal of neuroscience : the official journal of the Society for Neuroscience, Vol. 20, Issue 9, pp. 3182-90, (2000) (PubMed).
: "Characterization of the early molecular changes in the glomeruli of Cd151-/-mice highlights induction of mindin and MMP-10." in: Scientific reports, Vol. 7, Issue 1, pp. 15987 (PubMed).
Aliase für GDNF Antikörper
glial cell derived neurotrophic factor (Gdnf) Antikörperglial cell derived neurotrophic factor a (gdnfa) Antikörper
glial cell derived neurotrophic factor (GDNF) Antikörper
glial cell line derived neurotrophic factor (Gdnf) Antikörper
glial cell derived neurotrophic factor L homeolog (gdnf.L) Antikörper
AI385739 Antikörper
ATF1 Antikörper
ATF2 Antikörper
gdnf Antikörper
GDNF Antikörper
gdnfa Antikörper
hfb1-gdnf Antikörper
HFB1-GDNF Antikörper
HSCR3 Antikörper
Haben Sie etwas anderes gesucht?
- GDI2 Antikörper
- GDI1 Antikörper
- GDF9 Antikörper
- GDF7 Antikörper
- GDF6 Antikörper
- GDF5 Antikörper
- GDF3 Antikörper
- GDF2 Antikörper
- GDF15 Antikörper
- GDF11 Antikörper
- GDF10 Antikörper
- GDF1 Antikörper
- GDE1 Antikörper
- GDAP2 Antikörper
- GDAP1L1 Antikörper
- GDAP1 Antikörper
- GDA Antikörper
- GCS1 Antikörper
- GCP6 Antikörper
- GCP2 Antikörper
- GDP-Mannose Pyrophosphorylase B Antikörper
- GDPD1 Antikörper
- GDPD2 Antikörper
- GDPD3 Antikörper
- GDPD5 Antikörper
- Gelsolin Antikörper
- GEM Antikörper
- GEMC1 Antikörper
- GEMIN4 Antikörper
- GEMIN5 Antikörper
- GEMIN6 Antikörper
- GEMIN7 Antikörper
- GEMIN8 Antikörper
- Geminin Antikörper
- GEN1 Antikörper
- General Transcription Factor IIF, Polypeptide 2, 30kDa Antikörper
- Genome Polyprotein Antikörper
- Gephyrin Antikörper
- GFAP Antikörper
- GFER Antikörper



